Fragrance collection and tibial morphology
|
|||||||||||||||
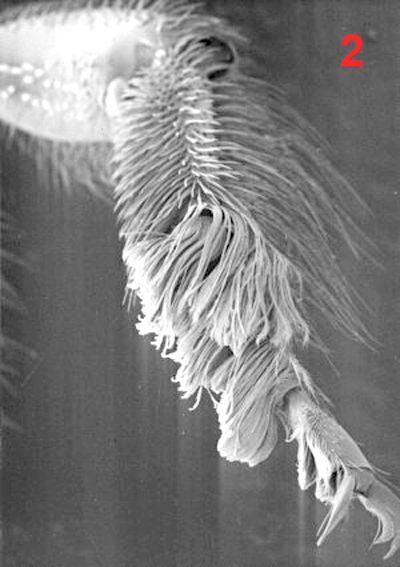
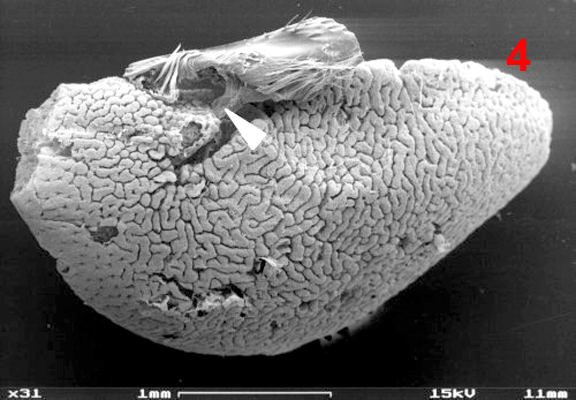
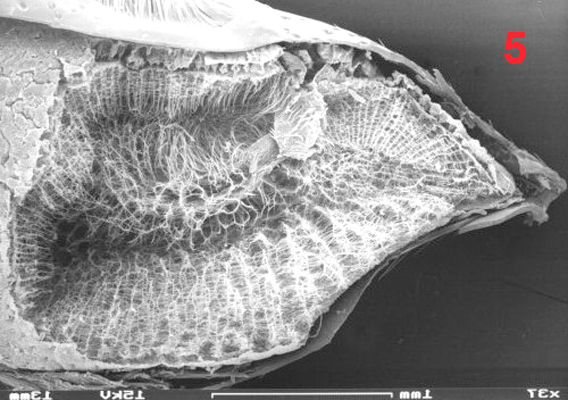
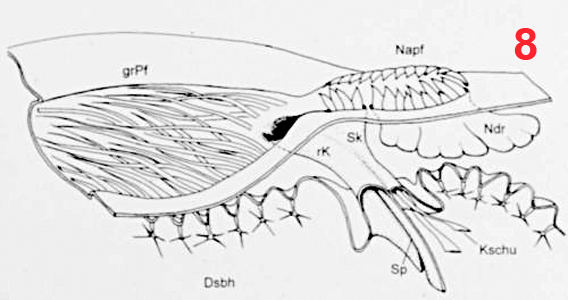
Male euglossine bees collect fragrances in a way analogous to that of 'enfleurage', an extraction process widely used in the perfume industry (Whitten et al. 1989, Eltz et al. 2007). Fist, the bees apply to the fragrant surface a drop of fatty lipids from their labial glands. The non-polar fragrance compounds become dissolved in the lipids and are mopped up together with them by the bees using their fore-tarsal brushes (1, 2, 3). Next, the mixture is stripped off onto the surface of the hind tibiae with the help of the mid-legs (1). Here, the liquid is absorbed and retained by the hind-tibial 'organ' (HTO), one of the most unusual structures found in bees. The anatomy of the HTO has been studied in great detail by Prof. Stefan Vogel in the 60th using histological methods (Vogel 1966). For my Diploma thesis (Eltz 1997) I examined the HTO of Euglossa tridentata, using both light microscopy and SEM. Apart from some details (perhaps originating from the fact that Vogel worked with E. cordata) my findings are in broad agreement with Vogels' work. I present my images and descriptions here because I know of no other published SEM shots of the tibial structures, and because Vogels papers as well as my thesis are in German. The HTO is essentially a cuticular sac. It represents a voluminous invagination of the exoskeleton that fills almost the entire tibia (4) and is connected to the outside by a short tube. The invaginated cuticle is heavily folded (4, 5) and produces dense stands of branched, tree-like hairs (6, 7), thus creating a sponge-like structure with a huge concealed surface area. The structurally most complicated part of the HTO is its interface with the outside world. In Euglossa the connecting tube actually consists of two subsections that are interconnected by a slit over their entire length. The two parts of the tube (one round and empty, the other narrow and filled with hairs) open into two separate zones of a hair-filled depression on the surface of the tibia (see schematic view in 8). The round part of the tube surfaces at the end of the larger, distal part of the depression which is covered by long and partly flattened hairs (9, 11). It is here where the mixture of lipids and fragrances is deposited during fragrance collection. Vogel hypothesized that the liquid is drawn into the sac by capillary forces via the round part of the tube. The narrow part of the tube surfaces towards the smaller, proximal part of the depression which is totally covered by layers of short scale-like hairs (9, 10, 11). Vogel speculated that the fragrances may eventually re-surface here, and this was later confirmed by tracer experiments with displaying males (of another species, Euglossa cognata, see Eltz et al. 2005). In addition to the described structures there is a second, much smaller cuticular invagination which originates from the proximal part of the depression (Ndr in 8). Most of the scale-like hairs covering the proximal depression actually grow out of this smaller invagination. In comparison to the main sac it is more heavily sclerotized. Also, it is lined by a layer of large cells inside the leg, suggesting that it may have secretory function. The tissue fringing the main sac is very thin and does not seem to have outstanding metabolic activity. Its function is yet unknown. My interpretation of the organ is that of a storage container. The sponge-like structure is likely to exert a strong capillary force that explains the bees' capability of storing lipid-fragrance mixtures for the very long periods of time observed (Eltz et al. 1999). There is little histological indication that fragrance components are absorbed from the sac, although this cannot be completely ruled out. Clearly, the organ is not completely impermeable for lipophilic chemicals, specifically the long chain carrier lipids originating from the cephalic labial glands. We have shown that those are selectively absorbed from the pouch and shuttled back to the labial gland, to be reused in consecutive bouts of volatile collection (Eltz et al. 2007). A similar fate may await some of the more lipoid exogenous fragrance compounds, but this has not been demonstrated. If any secretion of additional compounds takes place, this is likely to involve the tissue around the smaller sac that connects to the proximal section of the depression. Recently, Andreas Sager has completed his Diploma thesis entitled "Funktionsmorphologie der Duftakkumulation bei männlichen Prachtbienen (Apidae: Euglossini)". He compared the hind tibial morphology of representative species of all five euglossine genera by means of SEM. He found a surprising amount of variation concerning the container interface, size and structure of accessory invaginations, and the form of associated hairs. However, he also identified structural features that were common to all species, suggesting common functional principles. Vogel, S. (1966) Parfümsammelnde Bienen als Bestäuber von Orchidaceen und Gloxinia. Österr. Bot. Z. 113: 302-361. Whitten, W. M., Young, A. M., and Williams, N. H. (1989) Function of glandular secretions in fragrance collection by male euglossine bees. J. Chem. Ecol. 15: 1285-1295. Eltz, T., 1997. Zur Duftstoffbiologie neotropischer Prachtbienen (Apidae: Euglossini). Diplomarbeit, Universität Würzburg, 104 S. Eltz, T., Zimmermann, Y., Haftmann, J., Twele, R., Francke, W., Quezada-Euan, J. J. G. & Lunau, K., 2007. Enfleurage, lipid recycling, and the origin of perfume collection in orchid bees. Proceedings of the Royal Society B-Biological Sciences 274: 2843-2848. PDF Nature Research Highlight |
|
||||||||||||||

|

|
||||||||||||||
Lehrstuhl für Evolutionsökologie und Biodiversität der Tiere
Mitarbeiterhomepage PD Dr. Th. Eltz
RUB » Dekanat für Biologie und Biotechnologie » Evolutionsökologie und Biodiversität der Tiere - Mitarbeiterhomepage PD Dr. Th. Eltz